
C. David Allis
The Rockefeller University

Michael Grunstein
University of California, Los Angeles
For discoveries elucidating how gene expression is influenced by chemical modification of histones—the proteins that package DNA within chromosomes.
The 2018 Albert Lasker Basic Medical Research Award honors two scientists for discoveries that have elucidated how gene expression is influenced by chemical modification of histones, the proteins that package DNA within chromosomes. Through tour-de-force genetic studies in yeast, Michael Grunstein (University of California, Los Angeles) demonstrated that histones dramatically influence gene activity within living cells and laid the groundwork for understanding the pivotal role of particular amino acids in this process. C. David Allis (Rockefeller University) uncovered an enzyme that attaches a specific chemical group to a particular amino acid in histones, and this histone-modifying enzyme turned out to be an established gene co-activator whose biochemical capabilities had eluded researchers. Grunstein and Allis unveiled a previously hidden layer of gene control and broke open a new field.
Histone history
In the late 1800s, Albrecht Kossel discovered proteins called histones in goose blood cells. These abundant proteins, he showed, associate with nucleic acid to form a conglomerate called chromatin. Until the 1940s, many scientists thought that histones, not DNA, constitute the inherited material in eukaryotes, organisms whose cells contain nuclei.
By the 1960s, DNA had stolen the genetic-code limelight. Still, histones were plentiful and their partnership with the all-important genes intrigued investigators. Perhaps, evidence suggested, the proteins stifle the production of RNA from DNA, a process called transcription. In this view, stripping histones from eukaryotic DNA would allow the molecular apparatus that synthesizes RNA to adhere to its template and do its job.
In 1964, Vincent Allfrey (1921-2002), a biochemist at Rockefeller University, provided the first hints about events that might trigger such a process. Allfrey observed that histones from calf thymus nuclei that were adorned with acetyl chemical groups impair RNA synthesis poorly even though they bind DNA. On the basis of this finding, Allfrey made a bold hypothesis: The acetyl modifications create “presumably reversible changes in histone structure” that offer “a means of switching on or off RNA synthesis.”
Allfrey’s idea was provocative, but it rested largely on correlations between acetylation and gene activity. Rather than promoting transcription by fastening to histones, acetyl groups might instead affix to the proteins as a consequence of transcription. Although subsequent experiments from numerous investigators showed that histone acetylation characterizes active genes, the lack of cause-and-effect evidence left the idea without momentum.
In 1974, Roger Kornberg (Stanford University) proposed a theoretical model, now accepted, to explain how chromatin is organized. Two copies each of four distinct histone molecules combine to make a core around which DNA winds. Repeating units of this so-called nucleosome appear across the genome, like beads on a string.
By the late 1980s, elegant experiments by Yahli Lorch and Kornberg with purified cellular components suggested that nucleosomes can prevent the transcription machinery from accessing DNA and initiating RNA synthesis. But many scientists who studied transcription resisted the idea that nucleosomes inhibit gene activity. Because the evidence came from mixing reaction components in a test tube, no one knew whether these observations reflected events inside living cells. Furthermore, researchers had absorbed a paradigm from experiments on bacteria, which lack histones: Gene activators and repressors attach to specific spots in bacterial DNA to turn genes on and off. From this perspective, histones were not necessary to explain gene regulation. Instead, they were thought to function as inert structures that simply package DNA. Widespread thinking at the time held that nucleosomes were invisible to the transcriptional apparatus.
Histone-based gene control
In 1988, Grunstein catapulted the field forward. He genetically engineered yeast strains to shut down production of a particular histone, H4. This manipulation reduced the number of nucleosomes and boosted the amount of RNA synthesis from several test genes. The observation indicated that nucleosome loss in a living cell triggers gene activation and suggested that nucleosomes foil the transcription machinery’s ability to begin copying DNA into RNA. Grunstein had demonstrated that nucleosomes in intact eukaryotic cells do not serve merely as static spools that hold DNA; rather, they help regulate genes.
He discovered that one of the histone ends is needed to curb gene activity, and a specific amino acid in that N-terminal “tail”—a lysine that can be acetylated—plays a crucial role. Grunstein exploited the key lysine to identify molecules with which it collaborates, and showed that it interacts with a protein called SIR3, which was known to silence genes near chromosomal ends, where histones carry few acetyl groups. This finding, together with work from other labs, suggested a link between histone acetylation state and factors that regulate gene activity.
Grunstein went on to discover that the histone tail is required not just to quiet genes, but to activate them as well, and amino acid-substitution experiments indicated that acetylation of particular lysines is necessary for these effects. By counteracting the lysines’ positive charges, the acetyl groups presumably perturb chromatin structure in a way that promotes transcription (see Figure for one scenario).
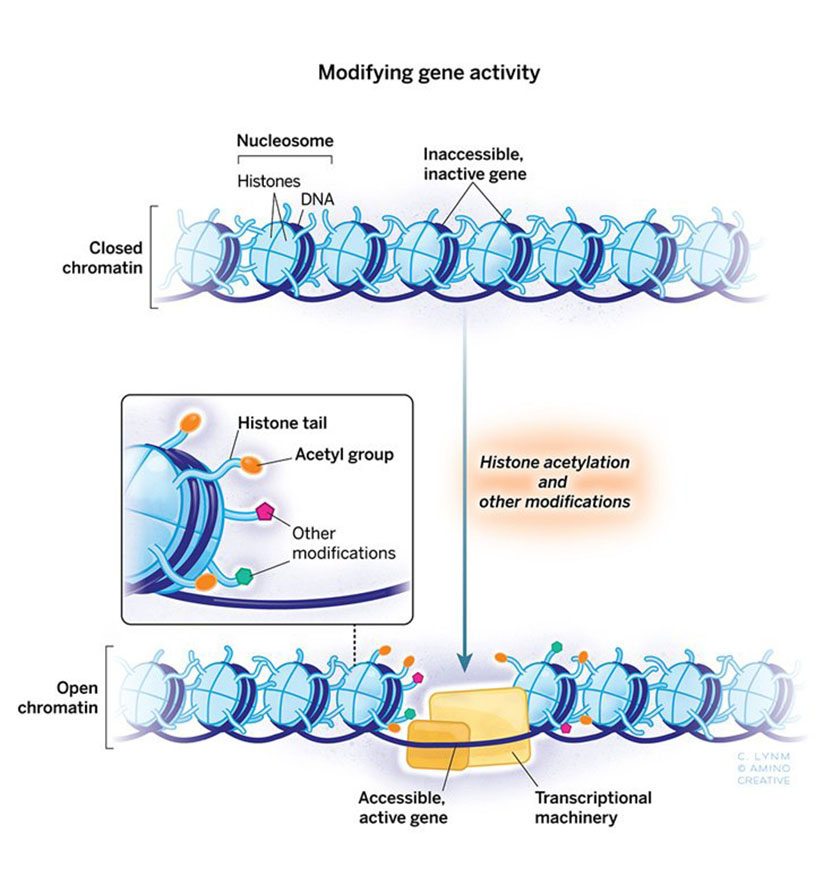
In one view of transcriptional activation by histone acetylation, unacetylated lysines, which carry a positive charge, allow the histones (light blue) to tightly bind DNA (dark blue), which carries a negative charge. Consequently, the transcriptional machinery cannot access the DNA, and genes remain inactive (top). Addition of acetyl groups (orange) to particular lysines in histone tails (box, middle) neutralizes the positive charge and loosens the nucleosome’s grip on DNA. This process allows the transcriptional machinery (yellow) to access the DNA, and genes are active (bottom). Other modifications (pink and green) affect transcription in different ways.
Modifying dogma
Although these experiments suggested that the acetylation state of histones contributes to gene control, acetylation per se had not been directly demonstrated in biochemical experiments. Allis had been seeking enzymes that perform this reaction, and in 1996, he reported a discovery that galvanized the field.
He was working with Tetrahymena thermophila, a single-celled pond-dwelling critter that has two nuclei, one of which percolates with transcription. Because histones in this nucleus are generously acetylated, Allis reasoned, they must contain an enzyme that adds acetyls to histones. He set out to find it.
Using 200 liters of Tetrahymena and an ingenious experimental system he devised to identify proteins that tack acetyl groups onto histones, he isolated such a protein. Sequence analysis of its gene revealed that it closely resembles a yeast protein called GCN5, a well-established co-activator of transcription. How GCN5 functions biochemically to turn on genes, however, was unknown. In a now-classic experiment, Allis showed that purified GCN5 was an enzyme that can acetylate particular lysines in histone tails—including some that Grunstein had pinpointed in his studies of transcriptional activation. The demonstration that a bona fide gene regulator acetylates histones provided another mechanistic tie between chromatin structure and gene control. A key question then arose: What property of GCN5 promotes gene activation? Is it the intrinsic catalytic activity of GCN5 or some other feature of the enzyme?
In 1998, Allis and, independently, Shelley Berger (University of Pennsylvania), showed that histone acetylation and gene activation in yeast fell if the catalytic activity of GCN5 was selectively abolished. This crucial demonstration cemented the principle that remodeling histone structure by enzymatic acetylation can determine where and when genes are triggered or subdued.
In the meantime, other researchers had been approaching the question from the other side—by pursuing enzymes that eliminate acetyl groups from histones. In 1990, Teruhiko Beppu (University of Tokyo) reported that a particular chemical inhibits removal of acetyl groups from histones in mammalian cells, and he began illuminating the biological effects exerted by this compound. Stuart Schreiber (Harvard University) followed up and found an enzyme that the inhibitor blocks. He showed that the enzyme bears sequence similarity to a yeast protein called RPD3, which previously had been shown to repress transcription of certain genes.
The field had now exploded. Enzymes could increase gene activity by adding acetyl groups to histones (GCN5) or decrease gene activity by removing them (RPD3). Moreover, by the end of 1996, Allis had demonstrated that a core component of the mammalian transcription machinery can acetylate histones, and a flurry of papers from other researchers were reporting similar findings that firmly extended the yeast and tetrahymena findings to mammals. Cells were bubbling with histone acetylase activity among proteins known to govern gene activity.
In 2000, Thomas Jenuwein (Max Planck Institute of Immunobiology and Epigenetics, Freiburg, Germany), in collaboration with Allis, discovered that an established transcriptional regulator was an enzyme that adds a different chemical group—a methyl—to a particular histone lysine. Others similarly connected gene activity with addition and subtraction of other chemical embellishments, including phosphate and ubiquitin moieties, to histones.
We now know that mistakes in histone modification touch a tremendous array of physiological pathways. Defects in the process underlie numerous inherited disorders of development—Kabuki syndrome and Rubinstein-Taybi syndrome, for instance—and these conditions affect multiple organ systems. Pharmaceutical companies are targeting the enzymes that attach or sever histone modifications with the aim of ameliorating human illnesses, especially certain forms of cancer.
By reshaping chromatin structure, chemical modifications of histones guide decisions about which stretches of DNA are read by conventional transcription activators and repressors. Grunstein and Allis have transformed our view of the histone proteins. Histones are not passive participants in DNA packaging, but rather, are key contributors to biological responses.
by Evelyn Strauss
Key publications of C. David Allis
Brownell, J.E., Zhou, J., Ranalli, T., Kobayashi, R., Edmondson, D.G., Roth, S.Y., and Allis, C.D. (1996) Tetrahymena histone acetyltransferase A: a homolog to yeast Gcn5p linking histone acetylation to gene activation. Cell. 84, 843-851.
Kuo, M.-H., Brownell, J.E., Sobel, R.E., Ranalli, T.A., Cook, R.G., Edmondson, D.G., Roth, S.Y., and Allis, C.D. (1996). Transcription-linked acetylation by Gcn5p of histones H3 and H4 at specific lysines. Nature. 383, 269-272.
Mizzen, C.A., Yang, X.J., Kokubo, T., Brownell, J.E., Bannister, A.J., Owen-Hughes, T., Workman, J., Wang, L., Berger, S.L., Kouzarides, T., Nakatani, Y., and Allis, C.D. (1996). The TAFII250 subunit of TFIID has histone acetyltransferase activity. Cell. 87, 1261-1270.
Kuo, M.-H., Zhou, J., Jambeck, P., Churchill, M., and Allis, C.D. (1998). Histone acetyltransferase activity of yeast Gcn5p is required for the activation of target genes in vivo. Genes Dev. 12, 627-639.
Rea, S., Eisenhaber, F., O’Carroll, D., Strahl, B., Opravil, S., Schmid, M., Mechtler, K., Pontig, C., Allis, C.D., and Jenuwein, T. (2000). Regulation of chromatin structure by site-specific histone methyltransferases. Nature. 406, 593-599.
Lewis, P.W., Mueller, M.M., Koletsky, M.S., Cordero, F., Lin, S., Banaszynski, L.A., Garcia, B.A., Muir, T.W., Becher, O.J., and Allis, C.D. (2013). Inhibition of PRC2 activity by gain-of-function mutations in pediatric gliobastoma. Science. 340, 857-861.
Maze, I., Noh, K.M., Soshnev, A.A., and Allis, C.D. (2014). Every amino acid matters: essential contributions of histone variants to mammalian development and disease. Nat. Rev. Genet. 15, 259-271.
Key publications of Michael Grunstein
Kayne, P.S., Kim, U.-J., Han, M., Mullen, J.R., Yoshizaki, F., and Grunstein, M. (1988). Extremely conserved histone H4 N-terminus is dispensable for growth but essential for repressing the silent mating loci in yeast. Cell. 55, 27-39.
Han, M., and Grunstein, M. (1988). Nucleosome loss activates yeast downstream promoters in vivo. Cell. 55, 1137-1145.
Johnson, L.M., Kayne, P.S., Kahn, E.S. and Grunstein, M. (1990). Genetic evidence for an interaction between SIR3 and histone H4 in the repression of the silent mating loci in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA. 87, 6286-6290.
Durrin, L.K., Mann, R.K., Kayne, P.S. and Grunstein, M. (1991). Yeast histone H4 N-terminal sequence is required for promoter activation in vivo. Cell. 65, 1023-1031.
Hecht, A., Laroche, T., Strahl-Bolsinger, S., Gasser, S., and Grunstein, M. (1995). Histone H3 and H4 N-termini interact with SIR3 and SIR4 proteins: a molecular model for the formation of heterochromatin in yeast. Cell. 80, 583-592.
Kurdistani, S.K., Tavazoie, S., and Grunstein, M. (2004). Mapping global histone acetylation patterns to gene expression. Cell. 117, 721-733.
Shahbazian, M.D., and Grunstein, M. (2007). Functions of site-specific histone acetylation and deacetylation. Annu. Rev. Biochem. 76, 75-100.

What Makes a Piece of Art or Science a Masterpiece?
Critics of art and philosophers of science have long wrestled with the question of what elevates a piece of art or a set of experiments to masterpiece status.
Award presentation by Richard Lifton
The very existence of life seems miraculous. The development from a single cell of a plant, an insect, or a human that each contains specialized cells and organs with radically different functions represents one of the great mysteries. We now understand that all life forms on earth use information encoded in DNA as the instruction manual for building an adult organism. Instructions are executed by copying segments of DNA, called genes, into RNA copies, that direct the production of the proteins that create a muscle cell, liver cell or neuron. This requires that different sets of genes are turned on, turned off, or tuned to the right levels in different cell types. Moreover, gene expression must also must be modulated to respond to periodic changes in the external environment.
Our first insights into how genes are selectively turned on and off came from studies of the bacterium E. coli by Jacob and Monod in the early 1960s. They established the paradigm that transcription factors- proteins that bind to specific DNA sequences- promote or inhibit the copying of DNA into RNA.
Acceptance remarks
Acceptance remarks, 2018 Lasker Awards Ceremony
What could be more “basic” than: 1) biomedical research—the belief that a mechanistic understanding of fundamental problems in biology will lead to translational advances, and 2) histone proteins—the proteins that serve to package our genome. I am a true believer in both. As an undergraduate I was exposed to doing “real experiments”, which, ironically, involved chasing elusive molecules in a cold room. This was a total turn on to me, and I quickly abandoned all thoughts about medical school. I wanted to tackle something at the heart of most biological processes—something in the nucleus of the cell—ideally working on gene regulation. I became intrigued with the biology of a pond water critter called Tetrahymena. A single-celled organism, Tetrahymena runs its life with two genomes partitioned in separate nuclei. One genome is kept in an active state, keeping the cell alive and well—perfect for exploring histone proteins. I was hooked.
The general view that histone modification was a critically important ‘switch’ for gene activation was championed more than 50 years ago by Vincent Allfrey who did all of his work at The Rockefeller University. But who put these chemical flags on; who took them off? This became the long-term goal of my lab, and Tetrahymena was going to be my secret weapon. I had no plan B, and nothing could have been farther from my mind than a Lasker Award.
Not everybody is cut out for science. Fortunately for me, Jim Brownell was. Jim was a graduate student in my lab some 25 years ago, and he is with us here today. Jim worked his magic in the cold room finding the gene switch that Allfrey had suggested might exist. Histones were doing more than just packaging of our genes—tthey were active participants in gene regulation, and Jim had found a protein switch that acted to turn them on. The implications of these findings were immediate and far-reaching. Histone modification was important; Allfrey had it right. Moreover, many of these proteins have proven to be druggable and some are FDA-approved with more on the horizon. These drugs are helping cancer patients lead healthier lives. Basic research and basic histones do matter.
To conclude, I never expected to win a Lasker Award, but I did always know that I had an amazing supporting cast. And as much as I like “histones”, I feel in love with a “herstone.” I thank my wife and family for all of their support. In 1997, Jim himself won a prize for his thesis and was interviewed after his prize ceremony. He commented, “Science is so funny. You are really famous among about a dozen people.” Jim said it right. We do what we do because it is interesting. It is a wonderful journey that I am privileged to take every day.
Congratulations to all of this year’s Lasker Awardees. Thank you all for this remarkable honor.
Acceptance remarks, 2018 Lasker Awards Ceremony
It is very humbling and a great honor to have been chosen for this award by the Lasker Jury and Foundation. I’m particularly pleased to share the honor with David Allis, whose contributions to the field of histone function have been pioneering.
And while David and I have the privilege of accepting the award, we do so in recognition that the Lasker not only awards the individual, but also the field of histone function. This area did not develop in a vacuum. We accept this award knowing that we have benefited from enormous advances in technology that began in the 1970s and that continue today. Our understanding of biology has always been linked to the progress of technology and that symbiosis has become so fruitful that the inventors could never have imagined the sophisticated extensions of their original findings
That is the history of this scientific discipline. It emphasizes that basic science of model microorganisms, funded mainly by the NIH, has led to the establishment of one of the most active fields in the biological and medical sciences and in the world-wide industry of biotechnology. Still, I believe that our understanding of the relevance of histone function is in its infancy. Histone associated proteins may prove themselves to be targets for novel pharmaceuticals. And technologies like gene editing may be able to reverse the defective histone modifications that are associated with certain disease states. David Allis will mention one such histone modification associated with pediatric brain cancer. There is even hope that reversals will one day be possible for certain neurological disorders, like RETT syndrome, given the finding that even advanced RETT symptoms are reversible in mouse models.
I consider myself extremely fortunate to have entered the field when I did. Was there a “eureka” moment? There was. It was when we replaced histone genes with certain mutant versions and saw these mutant histones incorporate into the chromatin of living yeast cells. Once we had a model system, our lab and others in the field could make changes, even in the most conserved regions of histone proteins, and ask new questions regarding novel histone functions. For instance, how can the same histones be involved both in gene activation and in gene repression?
I’ve struggled with the question of how to advise young scientists, since there is no one path to scientific discovery and it is often the discovery of the path itself that makes us so excited to be in Science. But at the expense of stating the obvious, the scientific questions we try to answer should be sufficiently bold so that we can invest ourselves with a focus, persistence and passion to address them.
Again, my thanks to the Lasker Foundation for this honor and to my family, friends and colleagues who have joined us here today.
