
Franz-Ulrich Hartl
Max Planck Institute of Biochemistry

Arthur L. Horwich
Yale University School of Medicine
For discoveries concerning the cell’s protein-folding machinery, exemplified by cage-like structures that convert newly made proteins into their biologically active forms.
The 2011 Albert Lasker Basic Medical Research Award honors two scientists for their discoveries concerning the cell’s protein-folding machinery, exemplified by cage-like structures that convert newly made proteins into their biologically active forms. With this work, Franz-Ulrich Hartl (Max Planck Institute of Biochemistry, Martinsried) and Arthur L. Horwich (Yale University School of Medicine) toppled traditional notions of how proteins fold inside cells and established new principles that operate from microbes to humans. This previously unexplored realm holds enormous importance for basic biology and biomedicine.
Protein folding is a vital process, as it converts linear amino acid chains into the three-dimensional forms that bestow the molecules’ unique activities. Greasy regions of new proteins, however, can grab one another and create useless globs. As proteins take shape, they bury these hydrophobic parts and expose hydrophilic, or water-loving, areas. Horwich and Hartl discovered that a special apparatus encases an unfolded protein and spurs folding by harnessing the energy of ATP, the small molecule that drives reactions inside cells.
However, larger, more complicated proteins than those studied by Anfinsen aren’t as self-sufficient and obliging. Furthermore, inside cells, protein concentrations are orders of magnitude higher than those that Anfinsen used, and as concentration rises, so does the risk of aggregation. Finally, new proteins in living creatures face a challenge that denatured, full-length proteins in a test tube circumvent: Because they gain a single amino acid at a time, portions of the growing chain can potentially stick to one another before the entire molecule is available to fold properly.
Unanticipated molecular caretaker
In the late 1980s, Hartl and Horwich were studying how proteins that are made in the cytoplasm enter mitochondria. Gottfried Schatz (University of Basel) and Walter Neupert (University of Munich) had shown that proteins are imported into mitochondria in a stretched-out state. Once inside, the amino acid sequences that targeted them to the mitochondria are removed, the proteins refold and, in many cases, assemble into multi-part structures before gaining enzymatic activity.
Horwich sought cellular machinery that participates in the mitochondrial-import process. He isolated yeast strains that fail to perform this essential task under certain conditions and then teamed up with Hartl (then at the University of Munich) to analyze the cells’ misbehavior. One of the mutants could transport proteins into mitochondria and clip them to the correct size — but the proteins lacked function, the team reported in 1989. This result suggested that the deviant yeast strain carries a damaged version of a component that normally facilitates protein folding or subsequent events, such as multi-molecular assembly, necessary for protein activity. The evidence strongly favored a block in folding, but assembly requires that a protein has achieved its proper form, so the experiments could not absolutely distinguish these possibilities.
The idea that some proteins need help to assemble had emerged from work on bacteria and chloroplasts. In 1972, Costa Georgopoulos (then at Stanford University) discovered that viral proteins fail to congregate into normal “head” structures within a strain of Escherichia coli that harbors particular genetic flaws. He subsequently tracked the defects to neighboring genes, groEL and groES. In 1980, R. John Ellis (University of Warwick) identified a chloroplast factor that physically interacts with newly made subunits of Rubisco, a key metabolic enzyme. He proposed that this Rubisco-binding protein promotes assembly of the enzyme. In 1988, these observations coalesced when Ellis and Georgopoulos sequenced the GroEL and Rubisco-binding protein genes and established their strong similarity. The researchers dubbed GroEL and the Rubisco-binding protein chaperonins, thus defining a subset of the chaperone family, whose members were thought to encourage macromolecular assembly by helping components avoid improper liaisons with themselves and others.
Genetic analysis revealed that Horwich and Hartl’s yeast mitochondrial protein was identical to the previously discovered heat shock protein 60 (Hsp60), whose production increases in response to heat — and that it resembles the chaperonins. Soon afterward, the researchers performed their dogma-defying experiment by testing whether folding per se — rather than assembly — depends on Hsp60. A protein that operates on its own relied on an Hsp60-based molecular machine to fold inside mitochondria, and ATP powered this reaction. Hartl and Horwich had thus demonstrated the existence of an ATP-driven ‘folding catalyst’ and revealed an unimagined piece of nature: Proteins imported into the mitochondria cannot refold spontaneously.
The mechanism takes shape
In the next advance, George Lorimer (Du Pont de Nemours & Co.) established that chaperonin activity could be studied using isolated components. He restored the enzymatic activity of denatured Rubisco in a test tube by adding to it purified GroEL and GroES in the presence of ATP. Horwich and Hartl set up a similar system to peer into the folding reaction. They showed, for instance, that GroEL (Hsp60’s bacterial counterpart) binds proteins in their relatively unstructured forms — and that addition of GroES and ATP provokes folding.
A tremendous amount of subsequent work, anchored by Hartl’s and Horwich’s ongoing independent investigations, provided key details about chaperonin action. For instance, the GroEL cavity, capped by the GroES lid, provides a cage where amino acid chains can fold, much as denatured proteins did in Anfinsen’s experiments, sequestered from unproductive interactions with other unfolded proteins. Furthermore, GroES cycles on and off GroEL — and GroEL sometimes ejects a protein that has not yet reached its active state. In that situation, the substrate protein rebinds GroEL for another try. These reports and others began to illuminate the sequence of events by which ATP orchestrates a cyclical reaction (see diagram) in which GroES and protein associate with alternating GroEL rings to enact folding.
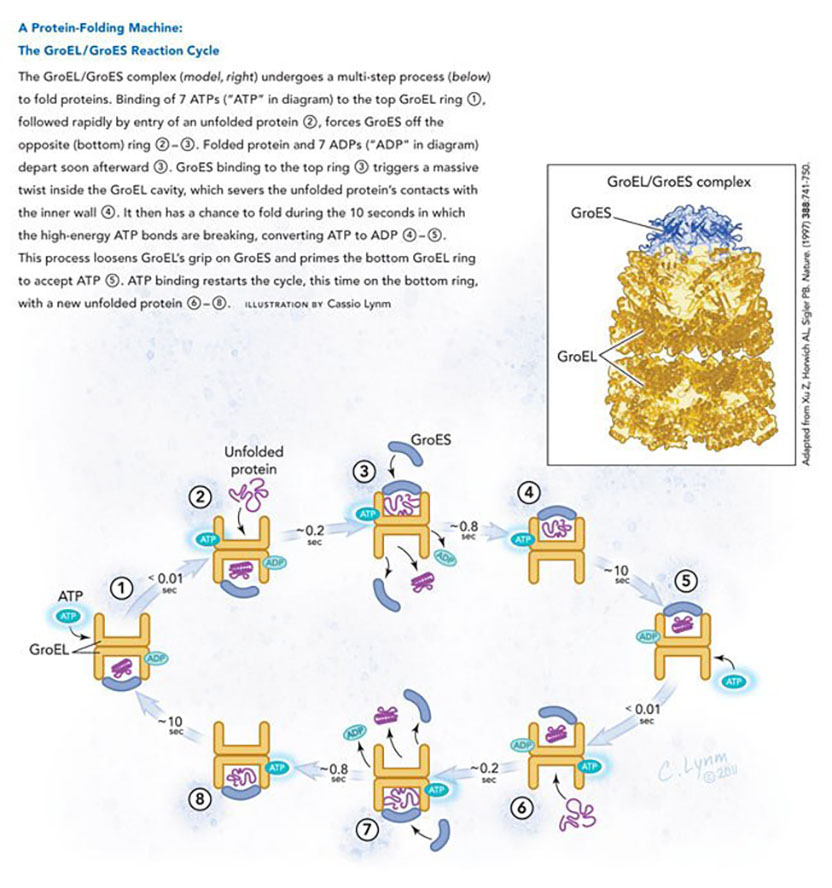
As these biochemical studies proceeded, Horwich and the late Paul Sigler, a Yale colleague, generated pictures at atomic resolution to elucidate details of the chaperonin reaction mechanism. They first discerned the structure of GroEL on its own, using X-ray crystallography.
Previous electron microscopic analysis had shown that the protein looks like two 7-fold symmetric donuts atop each other, and Horwich and Sigler found that oily amino acids point toward GroEL’s cavity at the donut’s opening. This observation suggested how GroEL selectively grabs unfolded proteins: through their hydrophobic portions. The researchers tested this prediction and others by probing whether specific amino acid alterations perturb key aspects of GroEL’s function. Changing hydrophobic elements to hydrophilic ones at particular spots inside the donut’s opening disrupted protein binding — and the amino acids that grasp protein also grip GroES, suggesting that protein and GroES compete for the same sites on GroEL. These results confirmed the team’s idea about how non-native proteins affix to the central channel and suggested how GroES attachment forces protein release into the cavity.
Three years later, Horwich and Sigler captured an X-ray image of the step after GroES caps the GroEL hole. The scientists then compared their snapshot of the combined molecules to pictures of the separate components. Upon binding to GroES and ADP, part of the GroEL donut enlarges and twists dramatically, thus ripping the unfolded protein from its connections inside GroEL and freeing it within the cavity. The contortion radically changes the internal environment — from greasy to watery — thus encouraging the unfolded protein to expose its hydrophilic regions and bury its hydrophobic ones. Additional analysis revealed how ATP binding and hydrolysis drives the folding reaction.
As the GroEL/Hsp60 story developed, a puzzle emerged. Hugh Pelham (Medical Research Council, Cambridge) had proposed that a different heat shock protein, Hsp70, also binds to hydrophobic surfaces to limit inappropriate associations among partially denatured proteins. Hartl wondered why cells contain two systems that serve similar purposes.
In 1992, Hartl established the concept of a relay in which each chaperone plays a distinct role. He demonstrated that the E. coli version of Hsp70 prevents premature folding of the growing amino acid chain; it then transfers the complete protein to GroEL, which promotes folding. He and Horwich had previously discovered that cage-like chaperonins distinct from those in mitochondria, chloroplasts, and bacteria also exist in the eukaryotic cytoplasm, and Hartl showed that similar sequential pathways operate there as well.
Certain medical conditions underscore the significance of these findings. When proteins aggregate, illnesses such as Alzheimer’s disease, Huntington’s disease, and amyotrophic lateral sclerosis can arise, and adjusting chaperone activity might provide therapeutic benefit. In addition, a particular Hsp60 mutation has been associated with hereditary spastic paraplegia, an illness in which the legs weaken and stiffen.
Hartl and Horwich unveiled a hitherto unknown process that enables proteins to reach their biological potential. Across the tree of life, the folding machines isolate young proteins and create a transformative moment. Then the devices send forth the mature molecules to join the hustle and bustle that makes cells what they are.
by Evelyn Strauss
Key publications of Franz-Ulrich Hartl
Cheng, M.Y., Hartl, F.U., Martin, J., Pollock, R.A., Kalousek, F., Neupert, W., Hallberg, E.M., Hallberg, R.L. and Horwich, A.L. (1989). Mitochondrial heat-shock protein hsp60 is essential for assembly of proteins imported into yeast mitochondria. Nature. 337, 620-625.
Ostermann, J., Horwich, A.L., Neupert, W., and Hartl, F.U. (1989). Protein folding in mitochondria requires complex formation with hsp60 and ATP hydrolysis. Nature. 341, 125-130.
Langer, T., Lu, C., Echols, H., Flanagan, J., Hayer, M.K., and Hartl, F.U. (1992). Successive action of DnaK, DnaJ and GroEL along the pathway of chaperone-mediated protein folding. Nature. 356, 683-689.
Langer, T., Pfeifer, G., Martin, J., Baumeister, W., and Hartl, F.U. (1992). Chaperonin-mediated protein folding: GroES binds to one end of the GroEL cylinder, which accommodates the protein substrate within its central cavity. EMBO J. 11, 4757-4765.
Frydman, J., Nimmesgern, E., Ohtsuka, K., and Hartl, F.U. (1994). Folding of nascent polypeptide chains in a high molecular mass assembly with molecular chaperones. Nature. 370, 111-117.
Hartl, F.U., Bracher, A., and Hayer-Hartl, M. (2011). Molecular chaperones in protein folding and proteostasis. Nature. 475, 324-332.
Key publications of Arthur Horwich
Cheng, M.Y., Hartl, F.U., Martin, J., Pollock, R.A., Kalousek, F., Neupert, W., Hallberg, E.M., Hallberg, R.L., and Horwich, A.L. (1989). Mitochondrial heat-shock protein hsp60 is essential for assembly of proteins imported into yeast mitochondria. Nature. 337, 620-625.
Ostermann, J., Horwich, A.L., Neupert, W., and Hartl, F.U. (1989). Protein folding in mitochondria requires complex formation with hsp60 and ATP hydrolysis. Nature. 341, 125-130.
Braig, K., Otwinowski, Z., Hegde, R., Boisvert, D.C., Joachimiak, A., Horwich, A.L., and Sigler, P.B. (1994). The crystal structure of the bacterial chaperonin GroEL at 2.8 A. Nature. 371, 578-86.
Fenton, W.A., Kashi, Y., Furtak, K., and Horwich, A.L. (1994). Residues in chaperonin GroEL required for polypeptide binding and release. Nature. 371, 614-619.
Rye, H.S., Burston, S.G., Fenton, W.A., Beechem, J.M., Xu, Z., Sigler, P.B., and Horwich, A.L. (1997). Distinct actions of cis and trans ATP within the double ring of the chaperonin GroEL. Nature. 388, 792-798.
Horwich, A.L. and Fenton, W.A. (2009). Chaperonin-mediated protein folding: using a central cavity to kinetically assist polypeptide chain folding. Q. Rev. Biophys. 42, 83-116.
Award presentation by Titia de Lange
The 19th-century philosopher Schoppenhauer said: “Talent hits a target no one else can hit; genius hits a target no one else can see.”
The target hit by the two basic scientists we honor today concerns the question of how proteins achieve their active state in the cell. At the time of their discoveries, Ulrich Hartl and Arthur Horwich were the only ones to see that this problem was unresolved. The rest of us were blinded by the false tenet that this issue had long been settled.
The misconception about how proteins fold into their native structure originated from key observations made some 30 years earlier, by Christian Anfinsen at NIH. Anfinsen, who shared the 1972 Nobel Prize in Chemistry for his discovery, had taken a purified active enzyme and denatured it so that its long chain of amino acids would unfold. He then diluted out the denaturant and discovered that the protein could spontaneously regain its enzymatic activity. The conclusion was clear: the amino acid sequence of the enzyme’s polypeptide chain is sufficient to specify its enzymatic activity, and since the enzymatic activity depends on the three dimensional structure of the protein, it followed that a chain of amino acids can fold correctly without addition of other proteins or ATP, nature’s energy currency.
Acceptance remarks

Acceptance remarks, 2011 Lasker Awards Ceremony
Receiving the Lasker Basic Medical Research Award is a tremendous honor and a great privilege. I am pleased to receive this award together with Art Horwich, with whom I share exciting memories of the early phase of discovery in the molecular chaperone field. I am very grateful to the Lasker Foundation and especially to the Lasker Jury for selecting us, and I wish to express my gratitude to the many people who have supported and ‘chaperoned’ me along the way, especially my wife and long-term collaborator Manajit.
To be recognized by a Lasker Award is of course the dream of every biochemist or molecular biologist. I am amazed that this dream has become a reality for me. But we should not forget that our true reward as scientists is the privilege of doing something that we enjoy so deeply and that doesn’t feel like work at all. I am well aware that our discoveries were only possible because many others who came before us laid the foundation. This, and a large quantity of good fortune, allowed us to make the surprising finding, in the late 1980s, that proteins, the molecules that drive almost every biological process, reach their proper shape (their ‘fold’) by the help of other proteins, which we call molecular chaperones. This was unexpected in light of the widely held view at the time that protein folding occurs spontaneously. It was my mentor Walter Neupert at Munich University who introduced me to Art. Walter supported us with his insight and provided the environment in which our collaboration could be successful. The mechanisms we eventually uncovered are beautiful in their simplicity: A large barrel-shaped protein complex, called chaperonin, functions like a mini-test tube in the cell, allowing immature protein molecules to fold, one at a time, while being shielded from the densely populated cellular milieu. As a result, new protein chains are protected from clumping together — just like the human chaperone prevents improper interactions between young people.
I grew up in a small village in the northern part of the Black Forest. Early on in high school, I developed a strong interest in biology and chemistry. In my second year of medical school at Heidelberg University, I was given the chance to work in the biochemistry lab. I quickly realized that this was what I wanted to do, rather than practicing medicine. Who knows how many lives I saved by that decision. In any case, I was hooked by research, despite the frustrations when experiments didn’t work. I remember vividly the Hungarian guest researcher who spent hours and hours in the cold room isolating proteins. When he emerged, apparently quite exhausted, he would invariably say “Biochemie ist kein Spaziergang” (“Biochemistry is not a walk in the park”). Owing to many fortunate circumstances, I can say that for me biochemistry became the path to a very exciting and rewarding life.
In recent years, research in a number of laboratories provided evidence for the medical relevance of molecular chaperones in preventing the formation of protein aggregates that cause presently incurable diseases like Alzheimer’s, Parkinson’s, or Huntington’s disease. Hopefully, one day it will be possible to harness the power of molecular chaperones for the treatment of these diseases. Should this happen, I can honestly say it was not planned, since for the most part I just wanted to find out a bit more about the fascinating workings of cells, without any application in mind. Unfortunately, science policy makers often do not realize the potential benefits of supporting such basic research. It is to the credit of organizations like the Lasker Foundation that the public is made aware of how scientific progress is most often achieved, namely through pure curiosity-driven research.
Finally, let me say what a great pleasure and honor it is to receive this award here in New York, the city in which my wife and I have spent some of the best years of our life and where we have so many wonderful friends.
Acceptance remarks, 2011 Lasker Awards Ceremony
I thank the Lasker Foundation and the jury for this enormous honor, which goes on beyond anything I could ever have imagined. There’s a wonderful comment of Sir Isaac Newton that I think reflects my life in science: “I know not what I may appear to the world, but to myself I seem to have been only like a boy playing on the sea-shore, and diverting myself in now and then finding a smoother pebble or a prettier shell than ordinary, whilst the great ocean of truth lay all undiscovered before me.” I feel ever so lucky that we picked up the mif4 mutant pebble that informed about what is a genuinely beautiful machine that Mother Nature uses to assist the folding of proteins in cells. I feel incredibly lucky not only to have been allowed to understand the general action of this machine but to have had the thrill to actually see it at near atomic resolution and ultimately, along with others, dissect how it works. Seeing GroEL on a graphics display for the first time in fall 1993 side by side with Zbyszek Otwinowski, who solved the structure and is here today, and the late Paul Sigler was a spiritual experience.
But I love science for all the moments between these once- or twice-in-a-lifetime moments. I love Newton’s “sea-shore” — the quiet contemplation of how something might work, the collaborative discussions with Ulrich and Ming (who is here today from Taiwan) in those early times, and with everyone in my lab every day, where I work at the bench side by side with everyone. And I love the personal dirtying of my own hands in my own experiments, where failure is the common outcome, but where I invest my being, as if a boy on the sea-shore.
We have recently directed our focus to the neurodegenerative paralyzing motor disease ALS, to a form caused by protein misfolding, and I hope that we or others are able to pick up the shell that will be the understanding of the basic mechanism by which a misfolded protein causes neurotoxicity. That might lead to a rationally designed therapy for this devastating condition. I once again thank you all for honoring me today.
Interview with Franz-Ulrich Hartl and Arthur L. Horwich
Video Credit: Susan Hadary
