
Max D. Cooper
Emory University School of Medicine

Jacques Miller
The Walter and Eliza Hall Institute of Medical Research
For their discovery of the two distinct classes of lymphocytes, B and T cells – a monumental achievement that provided the organizing principle of the adaptive immune system and launched the course of modern immunology
The 2019 Albert Lasker Basic Medical Research Award honors two scientists for discoveries that have launched the course of modern immunology. Max D. Cooper (Emory University School of Medicine) and Jacques Miller (Emeritus, The Walter and Eliza Hall Institute of Medical Research) identified two distinct classes of lymphocytes, B and T cells, a monumental achievement that provided the organizing principle of the adaptive immune system. This pioneering work has fueled a tremendous number of advances in basic and medical science, several of which have received previous recognition by Lasker Awards and Nobel Prizes, including those associated with monoclonal antibodies, generation of antibody diversity, MHC restriction for immune defense, antigen processing by dendritic cells, and checkpoint inhibition therapy for cancer.
When Miller began his research—around 1960—scientists had uncovered some features of the adaptive immune system, which protects our bodies from microbial invaders, underlies immunological memory, and distinguishes self from foreign tissue. They knew that antibodies, soluble proteins whose quantities surge after infection, perform jobs that differ from tasks that rely on live, intact cells such as rejection of transplanted grafts.
From vestigial to invaluable
Miller had no inherent interest in this useless body part, but observations about mouse lymphocytic leukemia drew him toward it when he began his Ph.D. work at the Institute of Cancer Research, London. A recently discovered virus caused this cancer when administered at birth, but not later, and preliminary experiments suggested that it targeted the thymus. To test whether it could multiply only in the newborn thymus, Miller removed the organ in newborn mice and then injected the virus.
He didn’t have a chance to find out the answer.
The pups remained healthy at first, but after weaning, they developed diarrhea and began wasting away. They were deficient in blood, lymph node, and spleen lymphocytes—and also in plasma cells, which produce antibody. The animals mounted a poor antibody response after bacterial immunization. Moreover, they failed to reject skin transplanted from unrelated mice and even rats, as would animals with an intact immune system. Two-thirds of the mice died prematurely.
Neonatal thymectomy was crippling immune activities, Miller concluded. Soon after publication of his initial results, other groups reported similar findings from studies on rats and rabbits. Miller subsequently established that the thymus is sometimes essential in adult animals too—to replenish the immune system if it is damaged by irradiation.
The thymus, he reasoned, might make cells that circulate and gain immune capabilities, poised to attack foreign threats throughout the body. To test this idea, he implanted thymuses into neonatally thymectomized mice and then introduced skin from different donors. The animals rejected grafts from unrelated individuals, but tolerated them from the strain that furnished the thymus. The thymus therefore, was supplying immunological function. Furthermore, in the recipient’s spleen—where lymphocytes multiply after stimulation— a significant fraction of dividing cells came from the donor thymus. As Miller had conjectured, cells from the thymus could migrate and mature.
Miller published the work in 1961 and 1962, and he presented his results at several meetings. Many scientists initially resisted his interpretations, including leading immunologists such as Peter Medawar (1915-1987) and MacFarlane Burnet (1899-1985). How could a disposable organ serve a vital purpose?
The strongest specific objection was that the lab in which Miller worked was a converted horse stable. Perhaps it was crawling with pathogens that put the animals in a stressed state, and the thymectomy pushed them into immunosuppression.
To address this possibility, Miller went to the U.S. National Institutes of Health, the only place in the world that had germ-free mice at the time. He repeated the experiments with those animals and confirmed that neonatally thymectomized mice accept foreign skin grafts. The immunological effects mapped to thymus removal, not to a pre-weakened immune system.
Dual lineage defense
In the meantime, Cooper, a pediatrician, had become interested in conditions that make children unusually susceptible to infections and, in 1963, he joined the laboratory of the late Robert A. Good at the University of Minnesota Medical School to explore his ideas. Some of the syndromes seemed to represent discrete, isolated segments of the adaptive immune system. For instance, many children with Bruton type agammaglobulinemia make no detectable antibody, yet can perform cell-mediated immune tasks. Conversely, children with other immunodeficiency conditions display the opposite physiological signature.
Such observations suggested the existence of two distinct arms of adaptive immunity in humans. By this time, however, the prevailing model, derived from Miller’s and other animal studies, suggested that a single pathway leads from thymus-generated lymphocytes to plasma cells. In this scenario, lymphocytes emerge from the thymus and disseminate to other tissues, such as the spleen, where stimulation with foreign substances triggers some of them to become antibody factories. The single-pathway model did not easily square with the clinical observations, which showed that cell-mediated activities could wither without wiping out antibody production. If lymphocytes give rise to plasma cells, it was not straightforward to explain how plasma cells and their products could abound when lymphocytes were scarce.
Results published far from the immunology mainstream pointed toward the chicken as a way to resolve this conundrum. In 1956, the late Bruce Glick (then a graduate student at Ohio State University) had reported in the Journal of Poultry Science that removal of a lymphoid organ—the bursa of Fabricius— soon after hatching could subdue antibody production. Perhaps, Cooper reasoned, he could use the chicken to simulate the apparent split reflected in the human diseases and tease apart potential contributions of the bursa and the thymus.
Cooper’s initial attempts to eliminate thymus function by removing the chicken thymus yielded no effects, and he speculated that newly hatched chickens might carry thymus-derived cells that had escaped from their birthplace before hatching, thus obscuring defects that should, theoretically, result from thymectomy. To address this possibility, he removed the thymus or the bursa of Fabricius from newborn chickens and then irradiated the animals to destroy residual immune cells that might have been floating around the body. Then he let the animals recover.
Irradiated chickens without a bursa did not produce antibodies after injection with bacteria or a foreign protein, and they lacked plasma cells. Their thymuses developed normally and lymphocytes flourished. In contrast, irradiated chickens with no thymus had few lymphocytes, but more than half made antibodies when stimulated. Subsequent studies showed that these animals could not reject skin grafts normally or execute other cell-mediated immune reactions.
In 1965 and 1966, Cooper published these findings and proposed that the bursa of Fabricius is essential for antibody production and the thymus is essential for cell-mediated immune responses. The names of bursa- and thymus-derived cells eventually shrunk to B- and T-cells, respectively. The two-pathway model provided a new lens through which to view human immunodeficiency diseases and brought fresh insight to a vast range of basic and clinical issues (see Figure).
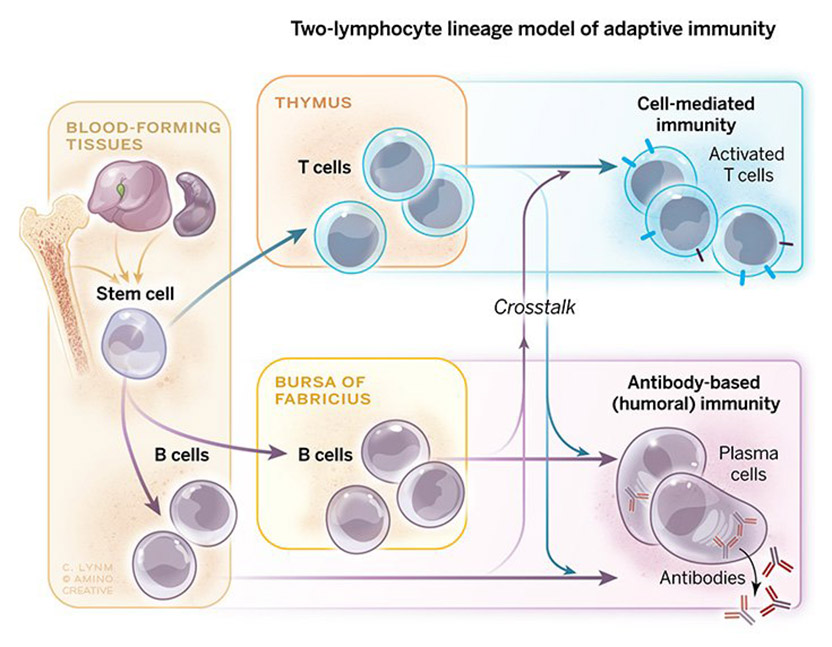
Dual-armed defense system
Stem cells from blood-forming tissues such as spleen, bone marrow, and fetal liver mature along the T-cell (top) or the B-cell (bottom) lineage to form, respectively, the cell-mediated or antibody-based components of the adaptive immune system. Along the way, signals from each type of developing cell prompt maturation and specialization of the other. In birds, the bursa of Fabricius hosts early stages of B-cell generation and antibody diversification, whereas in mammals, analogous steps occur in the bone marrow.
Illustration: Cassio Lynm / © Amino Creative
Crosstalk power
Even if these findings extended to mammals, which possess no obvious bursa equivalent, they raised perplexing questions. For instance, thymectomy plus irradiation of chickens softened antibody output—and neonatal thymectomies in mice stymied it. The split between the two pathways was not clean.
Miller (having joined the faculty at The Walter and Eliza Hall Institute of Medical Research) and his Ph.D. student, Graham Mitchell, decided to probe how the thymus might influence antibody production in mice. After obliterating thymus activity, they introduced thymus-derived cells, genetically distinct bone marrow-derived cells, or both before provoking antibody manufacture.
Neither cell type on its own reconstituted antibody production in the spleen. Only animals that received cells derived from the thymus and bone marrow achieved this feat. A collaboration was occurring between the two cell types.
Additional experiments revealed that the bone marrow-derived cells were the ones that spit out antibody, but only with assistance from the thymus-derived cells. Not only had Miller demonstrated that the dual system operates in mammals as well as birds, he also had exposed what we now know as helper T cells. This work ignited an explosion of interest in lymphocyte interactions.
Although mouse bone marrow contained cells that could give rise to antibody producers, the bone marrow might have received them from elsewhere, so the search for the mammalian bursa equivalent continued. In 1974, Cooper in collaboration with Martin Raff and John Owen (University College London) as well as independent investigators in Melbourne and Geneva, established that mammalian B cell precursors are generated in the blood-forming tissues—the liver in the fetus and the bone marrow after birth. This finding dovetailed with previous work on antibody development by Cooper. He had shown that two classes of antibody, IgM and IgG, each known for specific activities, come from a single precursor B cell that switches from IgM to IgG production.
Many laboratories subsequently elucidated the details of the system that Miller and Cooper unveiled, which operates in all jawed vertebrates. Cooper (at the University of Alabama, Birmingham, and then Emory University School of Medicine) went on to discover that jawless vertebrates—represented by the lamprey and hagfish—deploy a similarly organized scheme to recognize a vast array of foreign molecules, but with molecules that are structurally unrelated to the antibodies and receptors used by B and T cells.
By delineating the adaptive immune system’s two major branches, each of which performs distinct functions, Cooper and Miller opened a new era of cellular immunology. Virtually all fundamental discoveries in the field over the last 50 years can be traced to their pioneering work. Moreover, their historic findings have powered novel therapeutic strategies that have harnessed immune cells and their products to combat a vast range of illnesses—from cancer to autoimmune disorders to immunodeficiency conditions and far beyond.
by Evelyn Strauss
Key Publications of Max D. Cooper
Cooper, M.D., Peterson, R.D.A., and Good, R.A. (1965). Delineation of the thymic and bursal lymphoid systems in the chicken. Nature. 205, 143-146.
Kincade, P.W., Lawton, A.R., Bockman, D.E., and Cooper, M.D. (1970). Suppression of immunoglobulin G synthesis as a result of antibody-mediated suppression of immunoglobulin M synthesis. Proc. Natl. Acad. Sci. USA. 67, 1918-1925.
Cooper, M.D., Lawton, A.R., and Bockman, D.E. (1971). Agammaglobulinaemia with B lymphocytes. Specific defect of plasma-cell differentiation. Lancet. 2, 791-794.
Owen, J.J.T., Cooper, M.D., and Raff, M.C. (1974). In vitro generation of B lymphocytes in mouse foetal liver – a mammalian “bursa equivalent”. Nature. 249, 361-363.
Pancer, Z., Amemiya, C.T., Ehrhardt, G.R.A., Ceitlin, J., Gartland, G.L., and Cooper, M.D. (2004). Somatic diversification of variable lymphocyte receptors in the agnathan sea lamprey. Nature. 430, 174-180.
Cooper, M.D. (2015). The early history of B cells. Nat. Rev. Immunol. 15, 191-197.
Key Publications of Jacques F.A.P. Miller
Miller, J.F.A.P. (1961). Immunological function of the thymus. Lancet. 2, 748-749.
Miller, J.F.A.P. (1962). Effect of neonatal thymectomy on the immunological responsiveness of the mouse. Proc. Roy. Soc. 156B, 410-428.
Miller, J.F.A.P. (1962). Immunological significance of the thymus of the adult mouse. Nature. 195, 1318-1319.
Miller, J.F.A.P., and Mitchell, G.F. (1968). Cell to cell interaction in the immune response. I. Hemolysin-forming cells in neonatally thymectomized mice reconstituted with thymus or thoracic duct lymphocytes. J. Exp. Med. 138, 801-820.
Miller, J.F.A.P. (2011). The golden anniversary of the thymus. Nat. Rev. Immunol. 11, 489-495.
Miller, J. (2019). How the thymus shaped immunology and beyond. Immunol. Cell Biol. 97, 299-304.
Perspectives on B and T Lympocytes
Watts, G. (2011). Jacques Miller: immunologist who discovered role of the thymus. Lancet. 178, 1290.
Gitlin, A.D., and Nussenzweig, M.C. (2015). Fifty years of B lymphocytes. Nature. 517, 139-141.

Seurat’s Dots: A Shot Heard ‘Round the Art World
How the color theories of a chemist have ricocheted across centuries of artistic and scientific imagination.
Award presentation by Craig Thompson
In this ongoing era of molecular biology, it is easy to forget that the functional unit on which biology is built, is the cell. Working independently, the two exceptional scientists that we honor today discovered that the adaptive immune system is composed of two distinct cell types, B cells and T cells. Through their studies of lymphocyte development, Max Cooper and Jacques Miller have provided the framework on which modern immunology is built.
Miller and Cooper’s combined discoveries have also provided the foundation for most of today’s immune therapies. Monoclonal antibody production would not be possible without the antibody repertoire created and stored within the B cell lineage. One such antibody will be recognized with today’s Clinical Lasker Prize. Similarly, the success of T cell-based therapies, such as the recent development of CAR-T cells to treat cancer, depend on the immunologic properties that T cells acquire as they undergo lineage-specific development in the thymus.
Acceptance remarks
Acceptance remarks, 2019 Lasker Awards Ceremony
I am truly honored to receive the Albert Lasker Award with Jacques Miller for our discovery of the T and B cell lineages and their pivotal roles in cellular and humoral immunity.
While growing up in rural Mississippi, nothing could have seemed more remote than a career in biomedical immunology. My first childhood encounters with cellular immunity came while fishing and exploring nearby streams; every summer I developed severe poison ivy, known now as a classic cellular immune reaction. My first memorable encounter with humoral immunity came from receiving rabies vaccination with painfully escalating inflammatory lesions that followed the 14 daily injections of rabies virus grown in rabbit brain. (This was well before Jeff Ravetch showed that this “Arthus phenomenon” was triggered by interaction between the constant regions of antibodies and their receptors.)
Ralph Platou, my pediatrics chief at Tulane, encouraged me to pursue an academic career and helped me obtain further training at the Hospital for Sick Children in London. A growing interest in congenital immune deficiencies and allergic diseases led to an immunology and allergy fellowship at University of California in San Francisco. My new mentor requested that on the way there I learn the immunofluorescence microscopy technique for use in studying cell-mediated immunity in a keratoconjunctivitis model. John Holborow, a British immunologist, agreed to teach me the immunofluorescence technique, but when I stated why I wished to learn it, he gently informed me that it was only useful for studying humoral immunity. Embarrassed by my naivety, I vowed if nothing else I would learn the difference between humoral and cell-mediated immunity.
A realistic opportunity to fulfill this vow came later in Minnesota, when I joined Robert Good’s research group soon after Jacques’s discovery of the critical role of the thymus in immune system development. Parenthetically, leading immunologists at the time were still vigorously debating whether antibodies were actually responsible for cell-mediated immunity.
My “eureka moment” was actually a “eureka week” as the results unfolded from our experiments coupling irradiation of newly hatched chicks with removal of their thymus or bursa of Fabricius. The complete elimination of B lineage cells and their antibody products in bursectomized and irradiated chicks, together with their restoration by non-irradiated bursal cells, clearly delineated the bursa-dependent differentiation pathway from the thymus-dependent pathway that is responsible primarily for cellular immunity.
The pieces of the puzzle provided by these results together with information derived from studies of immune system development in immunodeficient patients and thymectomized mice, alongside those of bone marrow stem cells, allowed us to draw a provisional map of how the T and B lymphocyte lineages are derived from hematopoietic stem cells.
Over the following decades this basic organizing principle has proven to be true for immune system development in all living vertebrate species. It has been extensively amplified and elucidated through the work of many immunologists to yield better understanding and treatment of a variety of diseases, prime examples of which we will hear more about today.
Acceptance remarks, 2019 Lasker Awards Ceremony
I am extremely humbled, honored and delighted to receive the 2019 Lasker Basic Medical Research Award and to share this prestigious award with my long-time colleague Max Cooper. We never worked together, but I have met him on many occasions and have read his published work with great interest. We have also shared both the 1990 Sandoz Prize for Immunology and the 2018 Japan Prize for Medicine and Medicinal Science.
My curiosity for how the body responds to infection began as a child. Although born in France, I spent part of my childhood in China and Switzerland, and escaped with my parents to Australia from China in 1941, because of the Japanese threat during World War II. That was the year my very beautiful eldest sister died from tuberculosis. She had contracted it at a boarding school in 1936 and, although my younger sister and I often played with her, even when she was coughing blood stained sputum, we never developed the disease. Her doctor was overheard stating to my mother that nothing was known about how the body resisted infection. That statement, and the fact that I grew up when World War II was raging in Europe and Asia, made me decide to study Medicine, and thereafter to be involved in Medical Research.
After my residency as an intern at the Royal Prince Alfred Hospital in Sydney, Australia, I was awarded a Fellowship that enabled me in 1958 to study for a doctorate in London at the Cancer Research Institute. It was as a result of my studies on mouse lymphocytic leukemia, a cancer which in mice begins in the thymus before spreading elsewhere, that I made the serendipitous discovery of the immunological function of the thymus—a long-neglected organ. This stresses how important serendipity is in making really novel discoveries. A whole new world opened up before me and my work became more and more exciting, as happened subsequently during the identification of T and B cells, aided by my first PhD student Graham Mitchell at the Walter and Eliza Hall Institute of Medical Research in Melbourne.
It is still now very exciting to me, to see that the thymus, once believed to be a useless vestigial organ, populated with cells which in 1963 were considered by Nobel Laureate Sir Peter Medawar “as an evolutionary accident of no very great significance”, is producing T cells involved essentially across the entire spectrum of tissue physiology and pathology. These cells act not just in reactions considered to be bona fide immunological, but also, to cite some examples, in metabolism, in tissue repair, in dysbiosis and in pregnancy. I also find it most rewarding to see that basic research on thymus function, first published in one of my papers in 1961, and on T and B cells a few years later, has sown the seeds that spawned the new era of immunotherapy which can now claim a seat in the therapeutic pantheon of oncology, next to and perhaps about to supersede surgery, radiotherapy and chemotherapy.
Before I end, let me warmly thank the Lasker Foundation for celebrating basic medical research and for having chosen Max Cooper and myself for this prestigious award.
