
Elizabeth H. Blackburn
University of California, San Francisco

Carol W. Greider
Johns Hopkins University, School of Medicine

Jack W. Szostak
Harvard Medical School
For the prediction and discovery of telomerase, a remarkable RNA-containing enzyme that synthesizes the ends of chromosomes, protecting them and maintaining the integrity of the genome.
The 2006 Albert Lasker Award for Basic Medical Research honors three scientists who predicted and discovered telomerase, an enzyme that replenishes the ends of chromosomes. In so doing, they unearthed a biochemical reaction that guards cells against chromosome loss and identified the molecular machinery that performs this feat. The work resolved perplexing observations about chromosome termini and explained how cells copy their DNA extremities.
In the 1930s, scientists surmised that protective caps — telomeres — ensure the propagation of chromosomes during cell division and prevent them from inappropriately melding with one another. The physical nature of these structures — and how they are constructed — eluded researchers until Elizabeth Blackburn, Carol Greider, and Jack Szostak performed their groundbreaking investigations in the late 1970s and 1980s. Blackburn showed that simple repeated DNA sequences comprise chromosome ends and, with Szostak, established that these repeats stabilize chromosomes inside cells. Szostak and Blackburn predicted the existence of an enzyme that would add the sequences to chromosome termini.
The beginning of the ends
In the 1930s, Barbara McClintock (Lasker 1981) and Hermann Muller independently inferred that the natural termini of chromosomes display special characteristics. Unlike ends generated by DNA breakage, they don’t fuse with each other. Furthermore, only chromosomal fragments containing intact ends persist when a cell duplicates. A distinct structure must seal chromosomes and confer these properties, Muller reasoned. He dubbed chromosome termini “telomeres” from the Greek telos for end and meros for part or segment. However, no one knew what made telomeres different from randomly generated ends.
A second telomere-related conundrum arose after researchers deciphered how eukaryotic cells — those with nuclei — copy DNA. The enzyme that performs this reaction should be unable to fully replenish linear DNA due to a peculiarity of its mechanism; each round of replication should generate a molecule missing a few building blocks, called nucleotides, on the DNA’s end. As a result, linear chromosomes — which house genes in eukaryotes — would shorten every time a cell divides. In 1972, James Watson (Lasker 1960) speculated that organisms with linear chromosomes need a strategy to maintain chromosome tips, a theory that became known as the “end-replication problem.” In parallel, Alexey Olovnikov suggested that the gradual loss of chromosome ends would lead to cellular senescence, a dormant state in which cells remain alive but can no longer divide or perform their normal functions. Although scientists discussed possible solutions to these problems, they did not have ways to test their ideas.
The ends in sight
In the late 1970s, Blackburn wanted to determine the sequence of DNA at the ends of a eukaryotic chromosome. Joseph Gall, then at Yale University, had found that the ciliated protozoan Tetrahymena thermophila contains many DNA minichromosomes. Because the molecules are small but abundant, the number of ends relative to the rest of the DNA is large. This feature of Tetrahymena allowed Blackburn, working in Gall’s lab as a postdoctoral fellow, to gather enough ends to sequence. Each was composed of a six-nucleotide sequence (CCCCAA) that was repeated 20–70 times. Similar sequences turned out to reside in other ciliates, but no one knew whether this odd feature appeared in distantly related organisms.
In 1980, Blackburn, by then running her own lab at the University of California, Berkeley, presented her work at the Nucleic Acids Gordon Research Conference. After her talk, she spoke with Jack Szostak, a yeast geneticist from Harvard Medical School. They decided to add the Tetrahymena repeats to the ends of linear DNAs and test whether the resulting DNA would persist in budding yeast. Szostak knew that non-chromosomal linear DNAs in yeast normally insert themselves into chromosomes or are destroyed by cellular enzymes, presumably because they behave as if they result from random fractures. The Tetrahymena sequences provided the first hope for yeast to retain such linear DNAs. The experiment worked, despite the vast evolutionary distance between budding yeast and Tetrahymena. The Tetrahymena telomeres protected the linear yeast DNA, allowing it to pass reliably from one generation to the next.
The researchers then identified short, distinctive repeats on the ends of normal yeast chromosomes and showed that this yeast telomeric sequence was tacked on to Tetrahymena ends that were present on linear DNA in yeast. Because yeast added its characteristic sequence to Tetrahymena telomeres, telomeres must not serve as templates for additional telomeric sequences. This finding and the varied number of repeats led Blackburn and Szostak to speculate that an enzyme adds telomeric sequences to chromosome ends. Such an act would replenish the genetic material predicted to be whittled away by DNA replication. This idea differed radically from other suggestions scientists had proposed to solve the end-replication problem.
The means to the ends
Blackburn, soon joined by Greider, who was then a graduate student, started seeking the hypothetical enzyme by looking for a substance that could affix telomeric repeats to chromosome ends in the test tube. Because Tetrahymena contained so many telomeres, the organism should provide a rich source of enzymes that act on them, the researchers reasoned. They added the contents of Tetrahymena cells to a mixture of radioactive nucleotides and small DNA pieces composed of the Tetrahymena telomere repeats, hoping to see the tagged nucleotides attach to the repeats. On Christmas Day 1984, Greider discovered that the Tetrahymena contents generated precisely the DNA pattern predicted for an enzyme that added the six-nucleotide repeats one building block at a time. Blackburn and Greider named the enzyme telomere terminal transferase.
Next, they wanted to figure out exactly how the telomere sequence was determined. They postulated that each organism’s enzyme contained an RNA or DNA component that could serve as a template. An RNA-destroying enzyme obliterated the telomere terminal transferase activity, so Blackburn and Greider concluded that an RNA must play a crucial role. In a tour de force of biochemistry, Greider purified the enzyme, which they now called telomerase, and showed that it contained both an RNA and a protein subunit. In her own lab at Cold Spring Harbor Laboratory, she completed this work by isolating the RNA-encoding gene. It indeed carried a sequence that could specify the Tetrahymena telomere repeats. Furthermore, cleaving that sequence demolished telomerase activity. To prove that the internal template determines the sequence of the telomeric repeats, Blackburn altered the crucial sequence in the RNA component of telomerase; this perturbation resulted in production of telomeres that correspond to the new sequence. Later, Tom Cech (Lasker 1988) purified the protein portion of telomerase, which adds nucleotides one by one to the chromosome ends, according to instructions from the RNA component.
In the meantime, Szostak and his postdoctoral fellow Victoria Lundblad established that the inability to restore telomeres imperils the cell. They had been seeking yeast mutants that could not properly elongate telomeres. This scheme should identify genes that are crucial for telomerase function, they reasoned. Telomeres in such mutants would shrink, Szostak and Lundblad predicted, and strains harboring such defects would lose their chromosomes over many generations. At first, the yeast would grow normally, but as the genetic material disappeared, the microbes would stop dividing. The researchers found such a strain — and named the gene responsible for the defect EST1 for “ever shorter telomeres.” The approach subsequently led to the discovery of other proteins required for telomere stability, including telomerase’s core protein. Furthermore, the finding provided the first experimental support for the end-replication problem: As predicted, the inability to replenish telomeres caused the structures to dwindle as cells reproduced. Moreover, the work implied that cells unable to solve the end-replication problem eventually senesce.
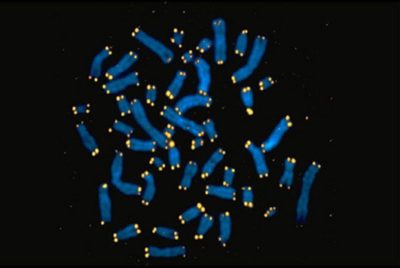
Visualizing chromosome tips. *For a full description of the figure, please see legend at the end of the essay.
All’s well that ends well
The work by Blackburn, Greider, and Szostak set the stage for discoveries about the role of telomerase in human cancer and aging. Like their Tetrahymena and yeast counterparts, human telomeres are composed of a particular simple DNA sequence, repeated various numbers of times. Sperm and eggs manufacture telomerase, but most adult cells don’t — and telomeres in most adult cells are shorter than those in sperm and eggs. Telomere attrition, at least in cells grown outside the body, leads to senescence.
Approximately 85 to 90 percent of human cancers reactivate telomerase (the rest maintain their telomeres through an alternative mechanism) and strong evidence suggests that the enzyme renders these cells able to proliferate uncontrollably by continually refreshing their telomeres. For example, adding the enzyme to certain human cells grown in culture dishes renders the cells immortal. Conversely, blocking the enzyme’s action in lab-grown cancer cells can inhibit their growth or kill them. Scientists are pursuing compounds that thwart telomerase as a potential strategy for fighting cancer. Several clinical trials of such drugs are now under way.
Along similar lines, telomere erosion during a person’s lifetime could curtail cell survival, thereby promoting age-related ailments. Evidence supporting this notion comes from studies of the rare human disease dyskeratosis congenita. One form of this illness arises from genetic defects in the RNA component of telomerase. Short telomeres limit the ability of certain tissues to replace themselves. As a result, the disease generates age-like conditions: It wipes out affected individuals’ bone marrow, predisposes them to a variety of human cancers, and gives them splotchy skin, ratty fingernails, and prematurely gray hair. The work on dyskeratosis congenita demonstrates that withered telomeres can accelerate physical deterioration.
Blackburn, Szostak, and Greider pursued basic questions of cell biology and enzymology to unveil mysteries that have huge implications for human health. The impact of their work is certain to extend long into the future.
by Evelyn Strauss
*Figure. Visualizing chromosome tips. Chromosomes (blue) carry telomeres (yellow) at their ends. The technique used for generating this image is a form of in situ hybridization, which was developed by Joseph Gall, this year’s winner of the Lasker Award for Special Achievement in Medical Science. To detect the telomeres, sequences that match the telomere repeats were tagged with fluorescent yellow molecules and applied to the chromosomes. The chromosomes were stained blue with a dye that binds DNA. [Credit: Peter M. Lansdorp, BC Cancer Research Centre, Vancouver, Canada]
Key publications of Elizabeth Blackburn
Blackburn, E.H. and Gall, J.G. (1978). A tandemly repeated sequence at the termini of the extrachromosomal ribosomal RNA genes in Tetrahymena. J. Mol. Biol. 120, 33–53.
Greider, C.W. and Blackburn, E.H. (1985). Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell. 43, 405–413.
Greider, C.W. and Blackburn, E.H (1987). A telomeric sequence in the RNA of Tetrahymena telomerase required for telomere repeat synthesis. Nature. 337, 331–337.
Yu, G.L, Bradley, J.D., Attardi, L.D., and Blackburn, E.H. (1990). In vivo alteration of telomere sequences and senescence caused by mutated Tetrahymena telomerase RNAs. Nature 344, 126–132.
McEachern, M.J. and Blackburn, E.H. (1995). Runaway telomere elongation cause by telomerase RNA mutations. Nature. 376, 403–409.
Kim, M.M., Rivera, M.A., Botchkina, I.L., Shalaby, R., Thor, A.D. and Blackburn, E.H. (2001). A low threshold level of expression of mutant-template telomerase RNA is sufficient to inhibit tumor cell growth. Proc. Natl. Acad. Sci. USA. 98, 7982–7987.
Key publications of Carol Greider
Greider, C.W. and Blackburn, E.H. (1985). Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell. 43, 405–413.
Greider, C.W. and Blackburn, E.H. (1987). The telomere terminal transferase of Tetrahymena is a ribonucleoprotein enzyme with two kinds of primer specificity. Cell. 51, 887–898.
Greider, C.W. and Blackburn, E.H (1987). A telomeric sequence in the RNA of Tetrahymena telomerase required for telomere repeat synthesis. Nature. 337, 331-337.
Allsopp, R.C., Vaziri, H., Patterson, C., Goldstein, S., Younglai, E.V., Futcher, C.W., Greider, C.W., and Harley, C.B. (1992). Telomere length predicts the replicative capacity of human fibroblasts. Proc. Natl. Acad. Sci. USA. 89, 10114–10118.
Prowse, K.R., Avilion, A.A., and Greider, C.W. (1993). Identification of a nonprocessive telomerase activity from mouse cells. Proc. Natl. Acad. Sci. USA. 90, 1493–1497.
Rudolph, K.L., Chang, S., Lee, H.W., Blasco, M., Gottlieb, G., Greider, C.W., and DePinho, R.A. (1999). Longevity, stress response, and cancer in aging telomerase deficient mice. Cell. 96, 701–716.
Key publications of Jack Szostak
Szostak, J.W. and Blackburn, E.H. (1982). Cloning yeast telomeres on linear plasmid vectors. Cell. 29, 245–255.
Murray, A.W. and Szostak, J.W. (1983). Construction of artificial chromosomes in yeast. Nature. 305, 189–193.
Blackburn, E.H. and Szostak, J.W. (1984). The molecular structure of centromeres and telomeres. Ann. Rev. Biochem. 53, 163–194.
Shampay, J., Szostak, J.W., and Blackburn, E.H. (1984). DNA sequences of telomeres maintained in yeast. Nature. 310, 154–157.
Dunn, B.L., Szauter, P., Pardue, M-L., Szostak, J.W. (1984). Transfer of telomere-adjacent sequences to linear plasmids by recombination. Cell. 39, 191–201.
Lundblad V. and Szostak, J.W. (1989). A mutant with a defect in telomere maintenance leads to senescence in yeast. Cell. 57, 633–643.
Award presentation by Joseph Goldstein
 The discovery of telomerase centers around four principal characters — a single-cell protozoan that swims in freshwater ponds, propelled by hairlike projections called cilia; a Tasmanian-born devourer of Amanda Cross mystery novels; a triathlete and competitive vaulter who performs handstands on the back of a speeding horse; and a 53-year-old virgin prizewinner.
The discovery of telomerase centers around four principal characters — a single-cell protozoan that swims in freshwater ponds, propelled by hairlike projections called cilia; a Tasmanian-born devourer of Amanda Cross mystery novels; a triathlete and competitive vaulter who performs handstands on the back of a speeding horse; and a 53-year-old virgin prizewinner.
So what is telomerase and why is it Lasker Prize-worthy? As you all know, life depends on the genetic instructions carried in the DNA of the genome.
Acceptance remarks

Acceptance remarks, 2006 Lasker Awards Ceremony
I feel greatly privileged to be recognized by a Lasker Award, and I want to convey my thanks and appreciation to the Selection Jury and to the Lasker Foundation for this very great honor.
Standing here today and thinking about the journey that has brought me here, I also feel a certain poignancy, because my mother, who is no longer alive, would have been particularly proud, and would have loved to be here. I remember the delight with which she would often tell me of how, when I was six years old, the teacher of a little rural grade school in the West of England, which I had been attending while my family was visiting from Australia, had said to her of me: “She will go far.”
This must have been gratifying indeed to hear for a parent who had had to live with my spine-chilling habit, in Australia, of picking up dangerous animals — poisonous jellyfish from the beach and stinging ants from twigs — and singing to them, behavior I thought perfectly natural because I loved animals. I was lucky to be given the circumstances that could transmute that childhood enthusiasm into a lifelong passion for doing science. I would wish everyone such good fortune.
I feel proud in turn of being part of a great tradition of biomedical research. I would like to see it stay strong and vibrant. To foster the best research and develop the best science policies, I want to say a couple of things that I believe we need to remember.
First, about research itself: It is important never to forget that science is as creative an endeavor as the humanities. Along with its more familiar aspects — like the necessary rigorous standards of proof — doing science is also letting the imagination be open to new ideas and lateral leaps that might at first seem outlandish. That means — as was true for our research that led to telomerase — having the freedom to do novel experiments at times, sometimes with obscure creatures. Joe Gall helped me appreciate this when I was a postdoctoral fellow with him. Because biology sometimes reveals its general principles through what may seem at first to be arcane and bizarre. All living things work fundamentally the same way when you get down to their molecules and cells; it is the extent and setting in which the various molecular processes are played out that differ from pond scum to humans, not their innermost workings.
The other thing I want to say is we certainly do not know where all our advances in understanding biology are headed: For a healthy science policy that serves society best, an environment of openness to available scientific evidence and freely shared and expounded ideas is crucial. We must surely keep alive a commitment to this. I feel very lucky to have been able to do science here in my adoptive country, the United States. But to realize the full promise of biological research we mustn’t fear new biological science, as sometimes seems to happen.
A scientific heroine of mine, Marie Curie, the discoverer of radium, said it well: “Nothing in life is to be feared. It is only to be understood.” Thank you.

Acceptance remarks, 2006 Lasker Awards Ceremony
It is indeed an honor to receive the Lasker Award. Working on telomeres and telomerase over the past 20 years has taken me on a ride though many disciplines in biology, from biochemistry to cellular senescence and aging, to recombination, the DNA damage response, and cancer. Today, we are delving into human genetics and stem cell biology. I have been very fortunate to have the opportunity to participate in these different research communities, to make new friends and collaborators, and to be exposed to new ideas and questions.
The most important lesson I have learned on this ride is that science is inherently a community activity. Ideas are created by thinking about your own experiments in the context of established knowledge and also thinking hard about other people’s experiments. When new ideas emerge they are discussed, critiqued, and modified. Often this process goes on unacknowledged. It is the exchange of ideas with the members of my laboratory that I find most exciting. Good ideas for the next most interesting experiments can come from many places; in my lab they almost always come from the students and postdocs who work with me. This lesson — that science is a community activity — is one of the most rewarding that I have learned.
I am grateful for the opportunity this award presents me to give back to the scientific community, by advocating the importance of fundamental, curiosity-driven research. The true value of high-profile awards is that it gives one the chance to explain publicly the nature of the scientific process and the importance of basic, non-applied work. Telomerase beautifully illustrates that you never know where medically relevant discoveries might come from. From studying Tetrahymena and yeast, we learn about cancer and stem cells. It has been a thrilling 20-year ride, I cannot wait to see where telomeres and telomerase will lead us in the next 20 years.

Acceptance remarks, 2006 Lasker Awards Ceremony
I would like to express my gratitude to the Lasker Foundation, and to Joe Goldstein and the other members of the jury, for this Award.
I am particularly happy to be recognized for my work on telomeres for several reasons — for one, the work was done so long ago that it has sometimes seemed to me to be almost forgotten. I am grateful to the jury for digging so deeply into the history of this field! Since then, I’ve worked in several rather different areas, but the experiment that Liz and I collaborated on, putting Tetrahymena telomeres into yeast, has always been one of my favorites.
I still remember discussing the idea with Liz at a Gordon Conference, and then the excitement when it actually worked and all of a sudden the door was opened to a whole series of new possibilities. To me, this experiment illustrates perfectly the value of talking to people who work in very different fields, the value of collaboration, and the value of, every now and then, putting a little money and effort into high risk, high payoff experiments.
It is a particular delight to be sharing this Award with Liz Blackburn, my collaborator from so long ago, and with Carol Greider, Liz’s former student who has done so much to develop the biochemistry of telomerase. When we did our work 20, 25 years ago, none of us could have anticipated the significance of the molecular biology of telomeres to aging and cancer. Our work was purely curiosity-driven basic science, and the biomedical significance emerged only later. I thank the Lasker Foundation once again for this recognition of our early work, and for their continued public support of basic research.
